Portrait of a Medicinal Plant** (Original title: Portraet einer Heilpflanze. Tropaeolum majus L. - die Kapuzinerkresse. Merkurstab 1995; 48:357-62. English by A. R. Meuss, FIL, MTA.)
Tropaeolum majus L. - Nasturtium
Ruth Mandera, Ulrich Meyer
Morphology
Generally known only as an ornamental plant, nasturtium was brought from
its native South America to Europe in the 17th Century. The Tropaeolum
genus derives from Chile and Peru where 80-90 species are found, their
distribution extending from the tropical rain forests to the snow line. The
slopes of the Peruvian Andes are said to be the natural habitat of nasturtium.
Alexander von Humboldt (1769-1859) pointed out that the flora growing
at different levels in the Andes actually reflect the plant cover of the whole
earth. Lower down, where the climate is warm and damp, the vegetation is
tropical, and in the cold atmosphere at high altitudes we find plants that also
grow in the polar region. Coming from the warmth (metabolic) pole of the
earth, close to the equator, the plant thus shows a tendency towards the cool
(neurosensory) pole.
The plant has made itself at home in Central Europe, showing surprisingly
lush growth and continuing to grow and flower throughout summer.
It likes a rich soil and is often found on compost heaps, for example. Oddly
enough, the stems do not come upright but stay close to the ground (Fig. 1).

Fig. 1.
Nasturtium
shoots stay close
to the ground.
Nasturtium has no lignifying, permanent support tissues and has to
depend on the support of hedges and fences if it is to reach any height. As a
rule, the fleshy shoots lie on the ground, producing adventitious roots if
covered with soil.
New leaves are continually developing at the shoot apex, with the first
signs of flower buds immediately apparent in the axils. The young leaves are
initially indented (Fig. 2).
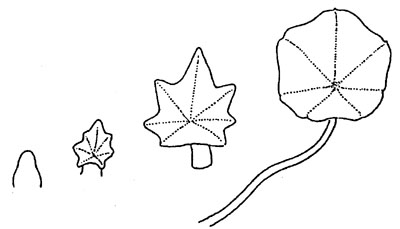
Fig. 2.
Development of
individual nasturtium
leaf.(1)
Even at this stage it is evident that the leaf is not
attached to the stalk at the margin, which is the usual way, but the lamina is
more or less centrally balanced on it (peltate leaf). As growth continues, the
fleshy stalks elongate, turning away from the earth towards the light, and
supporting the leaves, which now assume a horizontal position, from below.
These are now approximately circular (Fig. 3 and 4). Form and orientation are
remarkably like the floating leaves of water plants. Many other Tropaeolum
species have crenate or divided leaves, which tend to be smaller, and it
appears that large, juicy, rounded leaves are a special characteristic of
Tropaeolum majus.

Fig.3: Shoot
apex with young
leaves.

Fig. 4: Typical peltate
leaf.
An adequate water supply is vital in the hot summer months; even
temporary dryness will cause the soft leaves to hang flaccidly. The firm shape
can only be maintained if there is sufficient water. Hydathodes, cells active in
the excretion of water, are actually found in the leaf margins. After a cool
night in early summer, a droplet of water glitters at the end of every vein.
This marked relationship to water also explains why the vitality of these
plants is destroyed by even the slightest degree of frost in the fall. In no time
at all they turn yellowish and translucent, collapsing completely as the
freezing water expands and destroys their tissues.
In many European plants, vegetative growth terminates in an apical
inflorescence. The early stages of flower bud development frequently go
hand in hand with profound morphological and physiological changes in the
green parts of the plant.
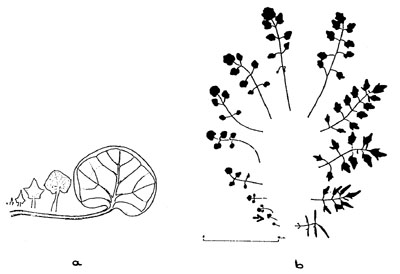
Fig. 5: Hairy bittercress (Cardamine hirsuta)
a) Evolution of ground leaf. b) Foliage leaf metamorphosis.(2)
Figure 5b shows the sequence
from first rosette leaf to the leaf
subtending the flower of hairy bittercress
(Cardamine hirsuta), Figure
5a the development of the second
leaf in its rosette (see arrow).
If we compare this with the
development of an individual nasturtium
leaf (Fig. 2), the similarity
is at first sight remarkable. Bittercress
quickly overcomes this "juvenile stage,"
however, and goes
into full expansion, division and
contraction of leaf forms before it
begins to flower. Nasturtium, on
the other hand, "blows" the juvenile
form up to considerable size and
repeats the same form all its life.
Flowering does not induce metamorphosis
of foliage leaves.
The phenomena show that nasturtium
has a strong connection to
the earth's surface and to water in its vegetative leaf sphere.
The large sulfur-yellow or orange-red flowers immediately reveal
another aspect of the plant. They grow singly from the leaf axils and stand
erect on long, fleshy stalks. The flowers show none of the innocence of a
buttercup that is open to the heavens. They are bilaterally symmetrical and face sideways, open to the animal that approaches them.
The colors of the five petals are brilliant, with the two upper ones
occasionally showing dark striations running to the center (Fig. 6).

Fig. 6: The two
upper petals of
a nasturtium
flower differ
slightly from
the other three.
The five sepals, which in other plants generally are as green as the foliage
leaves, have a yellow tinge in nasturtium. The most striking feature is a long
spur, which is also yellowy (Fig. 7). This has not developed from a sepal,
which is the usual way, but is an outgrowth of the flower stalk.

Fig. 7: Bud with spur.
Axis organs such as roots, stems and stalks are characteristically receptive
to earthly forces; they provide water and minerals for photosynthesis.
Here, the green stalk assumes color, creating an internal space with nectar
glands, and the characteristic spur of the nasturtium flower.
The distinction between leaf and flowering sphere is maintained, with
vegetative organs such as leaves and stem only subject to the flowering
impulse when very close to the flower (colored sepals, spur).
In their native habitat the flowers are pollinated by humming birds, their
beaks entering deep down into the spur to reach the sweet and also
somewhat hot (!) nectar. The impression is that the flower seeks to relate
closely to the bird in form and color.
The flower persists for about a week. The petals drop off soon after pollination
and the flower stalk bends over twice in a remarkable growth gesture,
going down so far that the fruits ripen beneath the leaves, close to the ground.
Nasturtium thus withdraws the fruit from the influence of light and warmth
(Fig. 8).

Fig. 8: Ripening fruit, showing stalk bending downward.
The fruit finally separates into three single-seeded parts, and the pulp,
originally spongy with a high water content, dries up to become cork-like
and shriveled.
Tropaeolum seeds contain an unusual fatty oil with an extraordinarily
high content of unsaturated fatty acids. Oils and fats from plants in tropical
regions characteristically contain saturated fatty acids (e.g. coconut and palm
kernel oil). Unsaturated fatty acids are mainly produced in the cooler regions
of the globe, examples being linseed and rape oil.(1)
The fatty acid spectrum of nasturtium oil reflects the "northern" character
of a mountain habitat and the cool, damp atmosphere in which the
seeds ripen.
Specific constituents
Morphologically, nasturtium deviates from the norm for flowering plants in
many respects. It also has a special constituent that is not to be found in just
any plant. (For mustard oil production in the cress family (Cruciferae), see
Rolf Dorka's "Zur Beziehung von Landschaft und Heilpflanze - Teucrium und Nasturtium als Tuberkulose-Heilmittel" (relationship between landscape and
medicinal plant - Teucrium and Nasturtium as tuberculosis medicines), Tycho
de Brahe-Jahrbuchfuer Goetheanismus, Niefem-Oeschelbronn, 1994.)
Nasturtium produces benzyl isothiocyanate, a volatile mustard oil with an
acrid, penetrant odor. This is hot to taste, and in highly concentrated, pure form
irritates the mucosa. Benzyl isothiocyanate may be regarded as a
thoroughly fiery, sulfurous compound.
The mustard oil is present throughout the plant but is not immediately
perceptible. It only develops its characteristic odor and taste when the tissue
is destroyed, e.g. by chewing a leaf or a flower. The plant "hides" the sulfurous qualities of the oil by binding it to sulfate (a salt-like or saline form of sulfur)
and sugar. The intact plant thus contains a "benzyl isothiocyanate sulfate
glycoside." This, in fact, makes the lipophilic benzyl isothiocyanate water-
soluble, so that it can be deposited in the vacuole. Metabolic end products are
characteristically "excreted" to the inside in vacuoles. Volatile oil plants often
let their material flow freely out into the atmosphere; nasturtium keeps its
mustard oil hidden deep inside the cell. When the plant tissue is damaged,
the enzyme myrosinase comes in contact with and is able to act on the glycosides
and "detonate the mustard oil bomb," as Zurich plant physiologist, Matile,
once put it. Nasturtium is able to control a highly sulfurous compound
such as benzyl isothiocyanate in saline form and store high concentrations of
it in all its organs.
The morphology shows tension between lush vegetative growth and powerful
flowering processes for the whole period of development. In the plant's
constituents the opposite qualities of Sal and Sulfur encounter each other.
Medicinal actions
Since the early 1950's, nasturtium has proved widely effective in the treatment of respiratory and urinary infections. Benzyl isothiocyanate inhibits or kills Gram positive and negative bacteria and fungi. To date, resistance has
hardly ever developed! It is interesting to note that the nasturtium action is
exclusively on the lungs, kidneys and bladder, organs that deal intensively
with the interplay of air and water.
Urinary tract infections frequently follow a chill, and this can be countered
with this "northern" yet thoroughly sulfurous plant.
Compared to phytotherapy, where it is given only by the oral route, nasturtium
plays a key role in the Wala acne preparations for topical use. Acne
vulgaris presents a paradox in skin metabolism. On one hand hyperkeratosis
produces blackheads. The masses of hardened keratin do not dissolve easily.
On the other hand sebaceous gland hyperactivity leads to seborrhea. The
excess sebum provides a nutrient base for bacteria which break it down into
fatty acids that cause skin irritation. Fatty acids are normally broken down
with the help of endogenous lipases in the human food metabolism.
With acne, we have a pathological degradation of fats by foreign organ-
isms on the skin, i.e. in the neurosensory system. Inflammatory efflorescences
go hand in hand with this.
Nasturtium may be said to be able to overcome the two fundamental
pathological processes in acne. It avoids all hardening processes and also has
a superb ability to control its mustard oil metabolism, keeping it in its proper
place (the vacuole).
Nasturtium in paintings
Nasturtium is a popular ornamental garden plant, but we know only of three
paintings that include it - Henri Matisse's La Ronde and Hannah Hoech's
Glaeser (Figs 9, 10). Henri Matisse (1869-1954) has painted at least two
versions of La Ronde, both in 1912. We shall limit ourselves to the one which
in our opinion is more characteristic. Hannah Hoech (1889-1978) was above
all esteemed as a Dadaist. Glaeser was painted in 1927.
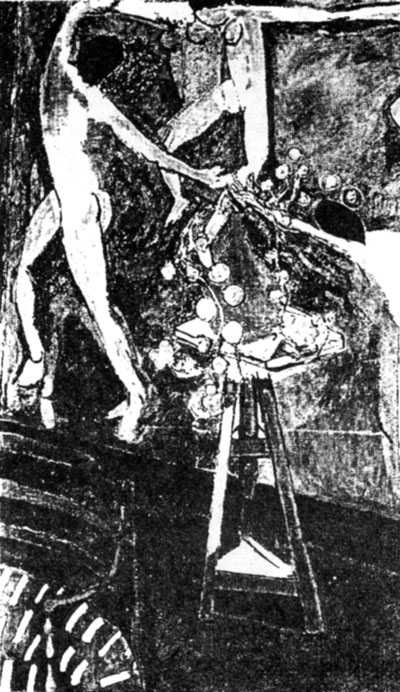
Fig. 9: La Ronde, by Henri Matisse.

Fig. 10; Glaeser, by Hannah Hoech.
It is perhaps not by chance that nasturtium appears so rarely in works of
art. It is not a cut flower to be portrayed at leisure - its "water form" will
survive only for a short time, even if put in water.
Both paintings show the round, peltate leaf. Matisse has the shoots
forming an approximate circle. The large, big-bellied vase is in deliberate
contrast placed on a small, square turntable. Behind the table dancers join in a
circle for their round dance.
Hannah Hoech also used the contrast between square and circle, with the
table top not only square but put at an angle. On the table are a number of
roundish vessels, marvelously showing the play of light and dark, with
circular reflections. A nasturtium leaf lies among the circles and vessels. Both
artists thus caught the characteristic gesture of the peltate leaf intuitively and
with seeming ease.
Ruth Mandera, Ph.D.
Ulrich Meyer, R.Ph.
References
1 Suchantke A. Die Zeitgestalt der Pflanze (1973). Goetheanistische Naturwissenschaft Band 2, Botanik. Stuttgart 1982.
2 Bockemuehl J. Bildebewegungen im Laubblattbereich hoeherer Pflanzen (1966).
Goetheanistische Naturwissenschaft Band 2, Botanik. Stuttgart 1982.
3 Errenst M. Die Waermeoffenheit von Fetten und Wachsen. Jahresbericht des Carl Gustav Carus-Institut, Niefem-Oeschelbronn 1992.
Photographs byJohannes Roth-Bernstein.